161.
Australian Institute of Marine Science 2023, Long-Term Monitoring Program Annual Summary Report of Coral Reef Condition 2022/23, Townsville.
162.
Emslie, M.J., Ceccarelli, D.M., Logan, M., Blandford, M.I., Bray, P., et al. 2024, Changing dynamics of Great Barrier Reef hard coral cover in the Anthropocene, Coral Reefs.
158.
Cheung, M.W.M., Hock, K., Skirving, W. and Mumby, P.J. 2018, Cumulative bleaching undermines systemic resilience of the Great Barrier Reef, Marine Ecology Progress Series 604(23): 263-268.
167.
Great Barrier Reef Marine Park Authority, Australian Institute of Marine Science and CSIRO 2024, Reef snapshot: summer 2023-24, Great Barrier Reef Marine Park Authority, Townsville.
174.
Morais, J., Tebbett, S.B., Morais, R.A. and Bellwood, D.R. 2023, Natural recovery of corals after severe disturbance, Ecology Letters 27(1): e14332.
185.
Thompson, A., Davidson, J., Logan, M. and Thompson, C. 2023, Marine Monitoring Program Annual Report for Inshore Coral Reef Monitoring: 2021–22. Report for the Great Barrier Reef Marine Park Authority, Townsville.
187.
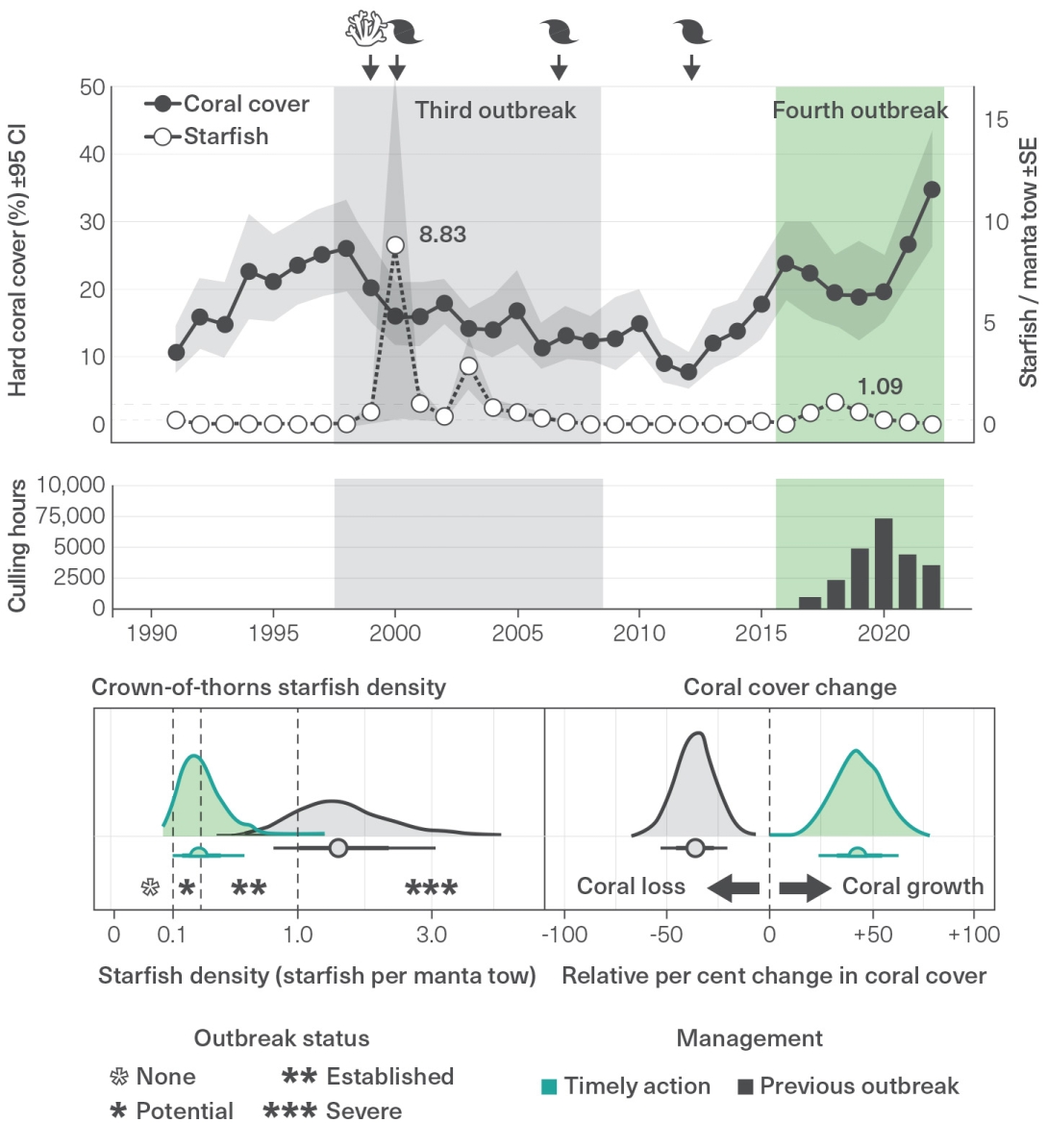
Matthews, S.A., Williamson, D.H., Beeden, R., Emslie, M.J., Abom, R.T.M., et al. 2024, Protecting Great Barrier Reef resilience through effective management of crown-of-thorns starfish outbreaks, PLoS One 19(4): e0298073.
-
311.
Hughes, T.P., Kerry, J.T., Álvarez-Noriega, M., Álvarez-Romero, J.G., Anderson, K.D., et al. 2017, Global warming and recurrent mass bleaching of corals, Nature 543(7645): 373-377.
370.
Wismer, S., Tebbett, S.B., Streit, R.P. and Bellwood, D.R. 2019, Young fishes persist despite coral loss on the Great Barrier Reef, Communications Biology 2(456): 1-7.
372.
Ceccarelli, D.M., Emslie, M.J. and Richards, Z.T. 2016, Post-disturbance stability of fish assemblages measured at coarse taxonomic resolution masks change at finer scales, PloS One 11(6): 1-22.
513.
Perrin, W.F. 2018, Spinner dolphin: Stenella longirostris, in Encyclopedia of Marine Mammals, pp. 925-928.
573.
Ortiz, J., Wolff, N.H., Anthony, K.R.N., Devlin, M., Lewis, S., et al. 2018, Impaired recovery of the Great Barrier Reef under cumulative stress, Science Advances 4(7): eaar6127.
751.
Pisapia, C., Hochberg, E.J. and Carpenter, R. 2019, Multi-decadal change in reef-scale production and calcification associated with recent disturbances on a Lizard Island reef flat, Frontiers in Marine Science 6(575): 1-10.
805.
Brandl, S.J., Rasher, D.B., Côté, I.M., Casey, J.M., Darling, E.S., et al. 2019, Coral reef ecosystem functioning: eight core processes and the role of biodiversity, Frontiers in Ecology and the Environment 17(8): 445-454.
874.
Tebbett, S.B., Morais, J. and Bellwood, D.R. 2022, Spatial patchiness in change, recruitment, and recovery on coral reefs at Lizard Island following consecutive bleaching events, Marine Environmental Research 173: 105537.
1072.
Morais, J., Morais, R.A., Tebbett, S.B., Pratchett, M.S., Bellwood, D.R. 2021, Dangerous demographics in post-bleach corals reveal boom-bust versus protracted declines, Scientific Reports 11(1): 18787.
1175.
Great Barrier Reef Marine Park Authority 2008, Cairns Area Plan of Management 1998, Great Barrier Reef Marine Park Authority, Townsville.
1176.
Great Barrier Reef Marine Park Authority and Office of Parliamentary Counsel 2018, Whitsundays Plan of Management 1998: Includes the 2017 amendment as in force 1 January 2018, Great Barrier Reef Marine Park Authority, Townsville.
1178.
Great Barrier Reef Marine Park Authority 2004, Hinchinbrook Plan of Management 2004, Great Barrier Reef Marine Park Authority, Townsville.
1604.
Hughes, T.P., Anderson, K.D., Connolly, S.R., Heron, S.F., Kerry, J.T., et al. 2018, Spatial and temporal patterns of mass bleaching of corals in the Anthropocene, Science 359(6371): 80-83.
1617.
Richardson, L.E., Graham, N.A.J., Pratchett, M.S., Eurich, J.G. and Hoey, A.S. 2018, Mass coral bleaching causes biotic homogenization of reef fish assemblages, Global Change Biology 24(7): 3117-3129.
1643.
Madin, J.S., Baird, A.H., Bridge, T.C.L., Connolly, S.R., Zawada, K.J.A., et al. 2018, Cumulative effects of cyclones and bleaching on coral cover and species richness at Lizard Island, Marine Ecology Progress Series 604: 263-268.
1644.
Richards, Z.T., Juszkiewicz, D.J., Hoggett, A. 2021, Spatio-temporal persistence of scleractinian coral species at Lizard Island, Great Barrier Reef, Coral Reefs 40(4): 1369-1378.
1650.
Wismer, S., Tebbett, S.B., Streit, R.P. and Bellwood, D.R. 2019, Spatial mismatch in fish and coral loss following 2016 mass coral bleaching, Science of the Total Environment 650: 1487-1498.
1700.
Baird, A.H., Álvarez-Noriega, M., Cumbo, V.R., Connolly, S.R., Dornelas, M., et al. 2018, Effects of tropical storms on the demography of reef corals, Marine Ecology Progress Series 606: 29-38.
1734.
Lewis, S.E., Bartley, R., Wilkinson, S.N., Bainbridge, Z.T., Henderson, A.E., et al. 2021, Land use change in the river basins of the Great Barrier Reef, 1860 to 2019: A foundation for understanding environmental history across the catchment to reef continuum, Marine Pollution Bulletin 166: 112193.
1888.
Pratchett, M.S., Caballes, C.F., Cvitanovic, C., Raymundo, M.L., Babcock, R.C., et al. 2021, Knowledge gaps in the biology, ecology, and management of the pacific crown-of-thorns sea star Acanthaster sp. on Australia's Great Barrier Reef, The Biological Bulletin 241(3): 330-346.
1984.
Great Barrier Reef Marine Park Authority 2024, Great Barrier Reef blueprint for climate resilience and adaptation, Reef Authority, Townsville.
2016.
Grafton, R.Q., Doyen, L., Béné, C., Borgomeo, E., Brooks, K., et al. 2019, Realizing resilience for decision-making, Nature Sustainability 2(10): 907-913.
2019.
Walker, B. and Salt, D. 2012, Resilience practice: building capacity to absorb disturbance and maintain function, 1st edn, Island Press, Washington, DC.
2028.
Thompson, A. A., Dolman, A. M. 2010, Coral bleaching: one disturbance too many for near-shore reefs of the Great Barrier Reef, Coral Reefs 29: 637-648.
2029.
Veron, J.E.N. 1995, Corals in space and time: the biogeography and evolution of the Scleractinia. University of New South Wales Press, Sydney, Australia.
2030.
Pandolfi, J.M. 2002, Coral community dynamics at multiple scales, Coral Reefs 21: 13-23.
2031.
McClanahan, T., Polunin, N., Done, T. 2002, Ecological states and the resilience of coral reefs, Conservation Ecology 6(2): 18.
2032.
Dornelas, M., Connolly, S. R., Hughes, T. P. 2006, Coral reef diversity refutes the neutral theory of biodiversity, Nature 440: 80-82.
2033.
Alvarez-Noriega, M., Madin, J.S., Baird, A.H., Dornelas, M. and Connolly, S.R. 2023, Disturbance-induced changes in population size structure promote coral biodiversity, The American Naturalist 202(5): 604-615.
2034.
Hughes, T.P., Graham, N.A.J., Jackson, J.B.C., Mumby, P.J. and Steneck, R.S. 2010, Rising to the challenge of sustaining coral reef resilience, Trends in Ecology & Evolution 25(11): 633-642.
2035.
Slater, J., Stokes, T., Great Barrier Reef Marine Park Authority 1997, Shoalwater Bay (Dugong) Plan of Management, Great Barrier Reef Marine Park Authority, Townsville.
2036.
Vanhatalo, J., Hosack, G.R. and Sweatman, H. 2017, Spatiotemporal modelling of crown‐of‐thorns starfish outbreaks on the Great Barrier Reef to inform control strategies, Journal of Applied Ecology 54(1): 188-197.
2037.
Great Barrier Reef Marine Park Authority, Australian Institute of Marine Science and CSIRO 2020, Reef snapshot: summer 2019-20, Great Barrier Reef Marine Park Authority, Townsville.
2038.
Dietzel, A., Connolly, S.R., Hughes, T.P. and Bode, M. 2021, The spatial footprint and patchiness of large-scale disturbances on coral reefs, Global Change Biology 27(19): 4825-4838.
2039.
Hughes, T.P., Kerry, J.T., Baird, A.H., Connolly, S.R., Dietzel, A., et al. 2018, Global warming transforms coral reef assemblages, Nature 556(7702): 492-496.
2040.
Hock, K., Wolff, N.H., Ortiz, J.C., Condie, S.A., Anthony, K.R.N., et al. 2017, Connectivity and systemic resilience of the Great Barrier Reef, PLoS Biology 15(11): e2003355.
2041.
Vercelloni, J., Liquet, B., Kennedy, E.V., Gonzalez-Rivero, M., Caley, M.J., et al. 2020, Forecasting intensifying disturbance effects on coral reefs, Global Change Biology 26(5): 2785-2797.
2042.
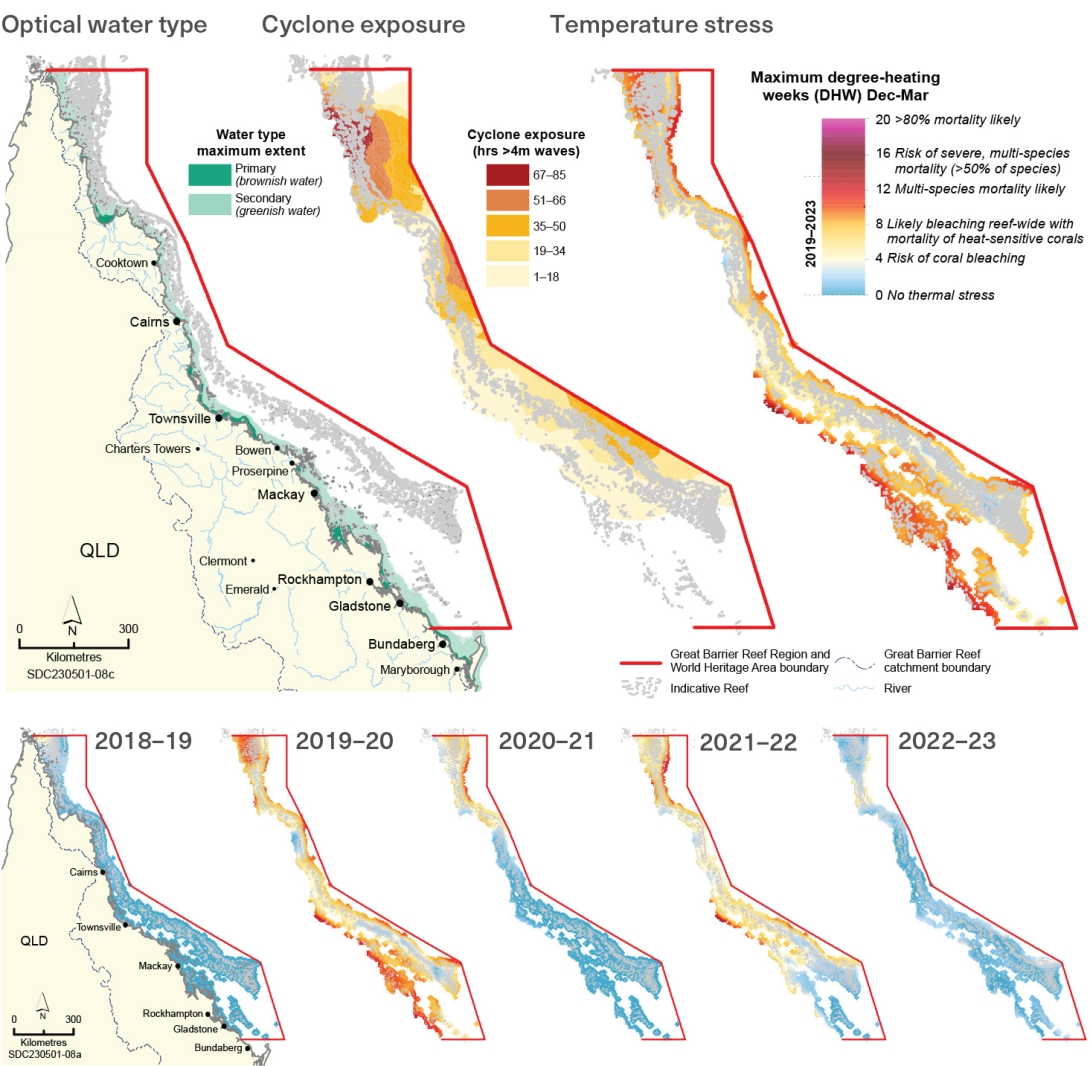
TropWater at James Cook University 2024, Optical water type data. Custom analysis provided to the Great Barrier Reef Marine Park Authority for use in the Great Barrier Reef Outlook Report 2024.
2043.
Puotinen, M. 2024, Custom analysis of cyclone exposure for the Reef Authority, Australian Institute of Marine Science.
2044.
NOAA Coral Reef Watch 2024, 'NOAA Coral Reef Watch Version 3.1 Daily Global 5km Satellite Coral Bleaching Degree Heating Week product for southern hemisphere summers 2018-19 to 2023-24. Custom analysis for the Great Barrier Reef Marine Park Authority. '. Unpublished report.
2045.
McWilliam, M., Pratchett, M.S., Hoogenboom, M.O. and Hughes, T.P. 2020, Deficits in functional trait diversity following recovery on coral reefs, Proceedings of the Royal Society B 287(1918): 20192628.
2046.
Dietzel, A., Bode, M., Connolly, S.R. and Hughes, T.P. 2020, Long-term shifts in the colony size structure of coral populations along the Great Barrier Reef, Proceedings of the Royal Society B: Biological Sciences 287(1936): 1-9.
2047.
Hughes, T.P., Kerry, J.T., Connolly, S.R., Alvarez-Romero, J.G., Eakin, C.M., et al. 2021, Emergent properties in the responses of tropical corals to recurrent climate extremes, Current Biology 31(23): 5393-5399.
2048.
Silverman, J., Schneider, K., Kline, D.I., Rivlin, T., Rivlin, A., Hamylton, S., Lazar, B., Erez, J., Caldeira, K. 2014, Community calcification in Lizard Island, Great Barrier Reef: a 33 year perspective, Geochimica et Cosmochimica Acta 144: 72-81.
-
2050.
Khan, J.A., Goatley, C.H.R., Brandl, S.J., Tebbett, S.B. and Bellwood, D.R. 2017, Shelter use by large reef fishes: long-term occupancy and the impacts of disturbance, Coral Reefs 36(4): 1123-1132.
2051.
Hoogenboom, M.O., Frank, G.E., Chase, T.J., Jurriaans, S., Álvarez-Noriega, M., et al. 2017, Environmental drivers of variation in bleaching severity of Acropora species during an extreme thermal anomaly, Frontiers in Marine Science 4(376): 1-16.
2052.
Álvarez-Noriega, M., Baird, A.H., Bridge, T.C., Dornelas, M., Fontoura, L., et al. 2018, Contrasting patterns of changes in abundance following a bleaching event between juvenile and adult scleractinian corals, Coral Reefs 37: 527-532.
2053.
Tebbett, S.B., Connolly, S.R. and Bellwood, D.R. 2023, Benthic composition changes on coral reefs at global scales, Nature Ecology & Evolution 7(1): 71-81.
2054.
Chow, C.F., Bolton, C., Boutros, N., Brambilla, V., Fontoura, L., et al. 2023, Coral settlement and recruitment are negatively related to reef fish trait diversity, Coral Reefs 42(2): 519-533.
2055.
Gonzalez-Rivero, M., Thompson, A., Johns, K., Ortiz, J., Kim, S., et al. 2023, Indicator Framework for the evaluation of the condition of coral reef habitats in the Great Barrier Reef: Methodological Documentation. Report prepared for the Great Barrier Reef Foundation, Australian Institute of Marine Science, Townsville.
2056.
Gonzalez-Rivero, M., Thompson, A., Johns, K., Ortiz, J., Kim, S., et al. 2023, Indicator Framework for the evaluation of the condition of coral reef habitats in the Great Barrier Reef: Introduction. Report prepared for the Great Barrier Reef Foundation, Australian Institute of Marine Science, Townsville.